3.1 作物产量
本期CropWatch通报监测期包含不同的作物生长季。南半球处于冬季作物(主要是小麦)收获期与夏季作物生长期之间;北半球冬季作物在通报撰写之前刚刚播种,高纬度区域冬季作物处于蛰伏期,最南部区域的作物当前处于加速生长期。夏季作物将在随后的几个月开始播种。
需要说明的是,表3.1中的环境因子指标数据,是本次监测时间段(2013年10月1日至2014年1月30日)数据与过去12年数据的平均距平所得。表3.1中的作物长势数据,是当前监测时间段数据与过去5年平均数据对比所得结果。
表3.1包含了一些变化较大的数据,如澳大利亚,乌克兰和波兰在监测期内的降雨量与之前12年平均水平相比分别偏低27%,25%和16%。同时,埃及本监测期内降雨量与近年平均相比也明显偏低,但由于埃及基本上都是灌溉作物,因此可以忽略其较少的降雨对作物生长的影响。降雨量明显多于近12年平均水平的国家包括乌兹别克斯坦(72%),墨西哥(48%),伊朗(42%),巴基斯坦(42%)和印度(52%),充足的降雨加速了这些国家的作物生长。作物生长环境因子全球分布图(图A.1至A.3)显示,主产国家中,极端旱情都发生在相邻国家的相邻区域,这符合自然分布的规律。但更加明显的异常可能会出现在主产国范围之外的其它国家。
监测期内,北美(美国气温偏低1℃,加拿大气温偏低1.3℃)和东南亚地区(泰国气温偏低1℃)气温较近12年平均偏低。欧洲从西伯利亚-俄罗斯至德国,气温与近12年平均相比,分别偏高1.7℃和1.2℃。阿根廷气温较近年平均偏高1℃,较高的温度导致阿根廷在夏季作物生长初期发生旱情。现在还不能确定欧洲的高温对冬小麦产量的影响程度,因为2月至收获期的天气状况对欧洲冬小麦最终产量仍然存在较大影响。
基于遥感的光合有效辐射数据与降雨和温度变化幅度相比要小,而且也没有极值出现。尽管如此,在中国、加拿大、土耳其和尼日利亚也出现了与近12年平均水平偏高幅度较大的情况(偏高6%-8%)。墨西哥和菲律宾由于降雨过多,光合有效辐射分别降低3%和4%。在墨西哥,水分是作物生长受限的一个主要的制约因素,充足的降水将会抵消光合有效辐射减少带来的负面影响。在菲律宾,台风“海燕”的巨大破坏作用以及寡照天气,使作物减产。
表3.1 全球粮食主产国环境因子和作物指数分别与过去5年和12年参考值的距平
| 环境因子和过去12年 | 作物指数和过去5年平均值的距平(2008-2013) | |||||
| 平均值的距平(2001-2013) | ||||||
| 累积降雨(%) | 平均气温(°C) | 累积PAR(%) | 累积潜在生物量(%) | 未播种的可耕地面积(%) | 最佳植被状况指数 | |
| 阿根廷 | 5 | 1 | 0.1 | -1 | 0.7 | -0.05 |
| 澳大利亚 | -27 | 0.3 | 3 | 3 | 9.2 | 0.01 |
| 孟加拉国 | 11 | -0.5 | -0.5 | 33 | -0.2 | 0.06 |
| 巴西 | -1 | 0.2 | -0.4 | 2 | -0.4 | 0.01 |
| 柬埔寨 | 5 | -0.8 | 5 | 8 | 0.5 | -0.01 |
| 加拿大 | 8 | -1.3 | 6 | -2 | 10.7 | 0.01 |
| 中国 | 19 | 0.5 | 8 | 21 | -3.3 | 0.03 |
| 埃及 | -24 | 0.2 | 3 | 26 | -1 | 0.05 |
| 埃塞俄比亚 | 28 | 0.3 | 0.2 | 16 | -4.3 | 0.01 |
| 法国 | -3 | 0.8 | 0.1 | 4 | -2 | 0.07 |
| 德国 | 5 | 1.2 | -0.4 | 8 | -10.7 | 0.11 |
| 印度 | 52 | -0.3 | 1 | 33 | -3 | 0.11 |
| 印尼 | -2 | -0.2 | -2 | -2 | 0.4 | -0.01 |
| 伊朗 | 42 | -0.4 | 2 | 19 | -2.8 | 0.06 |
| 哈萨克斯坦 | 25 | 2 | 4 | 17 | -1.9 | 0.23 |
| 墨西哥 | 48 | -0.1 | -3 | 31 | -4.3 | 0.04 |
| 缅甸 | 27 | -0.2 | 2 | 4 | -4.8 | 0.01 |
| 尼日利亚 | 23 | 0.3 | 6 | 9 | 1.6 | -0.01 |
| 巴基斯坦 | 42 | -0.1 | 3 | 14 | -3.5 | 0.04 |
| 菲律宾 | 11 | -0.3 | -4 | -0.2 | -0.4 | 0 |
| 波兰 | -16 | 1.2 | 4 | -7 | -24.2 | 0.09 |
| 罗马尼亚 | -8 | 1.2 | 2 | -6 | -12.5 | 0.1 |
| 俄罗斯 | -1 | 1.7 | 4 | 6 | 11.8 | 0.12 |
| 南非 | -3 | 0 | 3 | 0 | 3 | -0.03 |
| 泰国 | 13 | -1 | 5 | 6 | 0.6 | 0.02 |
| 土耳其 | -6 | -0.2 | 7 | -9 | 18.9 | -0.03 |
| 英国 | 42 | 0.7 | 1 | 10 | -3 | 0.09 |
| 乌克兰 | -25 | 0.7 | -1 | -11 | -30.5 | 0.04 |
| 美国 | -4 | -1.4 | 3 | -2 | 5.9 | 0.05 |
| 乌兹别克斯坦 | 72 | 0.5 | 4 | 52 | -8.6 | 0.01 |
| 越南 | -1 | -0.6 | 1 | -7 | -1.5 | -0.01 |
注:除了温度用用摄氏度表示之外,其他环境因子的距平用相对百分比来表示。0表示和过去平均比没有变化;相对距平用如下公式表示(C-R)/R*100C 表示当前季度的值,R表示过去的五年(5YA)2008年10月-2013年1月或者十二年(12YA)2001年10月-2013年1月同期的平均值。VCI为最佳植被指数。
潜在生物量与参考值间的距平(图3.1)可能由高于正常水平的温度或降雨导致的。降雨量对于温暖以及半干旱气候贡献相对最为显著;降雨量高于平均水平较多的区域,潜在生物量增加明显,其中:墨西哥增加31%,印度增加33%,乌兹别克斯坦增加52%。温度的作用在降雨为非限制条件或者灌溉区域较为显著,例如,中国潜在生物量增加21%。正如前面提到的,由于波兰降雨减少7%,罗马尼亚降雨减少6%,所以这两个国家的潜在生物量减少最大。土耳其和越南则都经历了相对轻微的干旱和低温,因此,这两个国家的潜在生物量分别减少9%和7%。
土耳其的未种植耕地比率增长(18.9%)是所有主产国中最大的,表明未种植的耕地急剧增加。尽管很难去评定背后的原因,但在3.2节国家详细分析中,基于潜在生物量分布图和VHI曲线清晰的呈现出了受影响的区域。其它未种植耕地比率大幅增加的国家中,美国和加拿大是由于长时间的低温所致。俄罗斯则未种植耕地比例增幅也高达11.8%,虽然很难解释其增加的原因,但俄罗斯国内公布的消息还是可以证实这一点是确实存在的。
监测时间段内最佳植被状态指数与近12年平均距平变化的特点在绝大多数情况下与环境指标变化趋势一致。哈萨克斯坦受降雨增加25%和温度偏高2°C的影响,最佳植被状态指数增幅最大,增加0.23。其次为一些欧洲国家,较高的温度导致最佳植被状态指数增加,例如,俄罗斯增加0.12,德国增加0.11。报告中,最佳植被状态指数降低最多的为阿根廷和南非,降幅分别为0.5和0.3(图3.2)。
图3.1 全球各国(包括大国的省州级别)累积潜在生物量与过去12年(2001-2013)的距平,单位百分比(%)
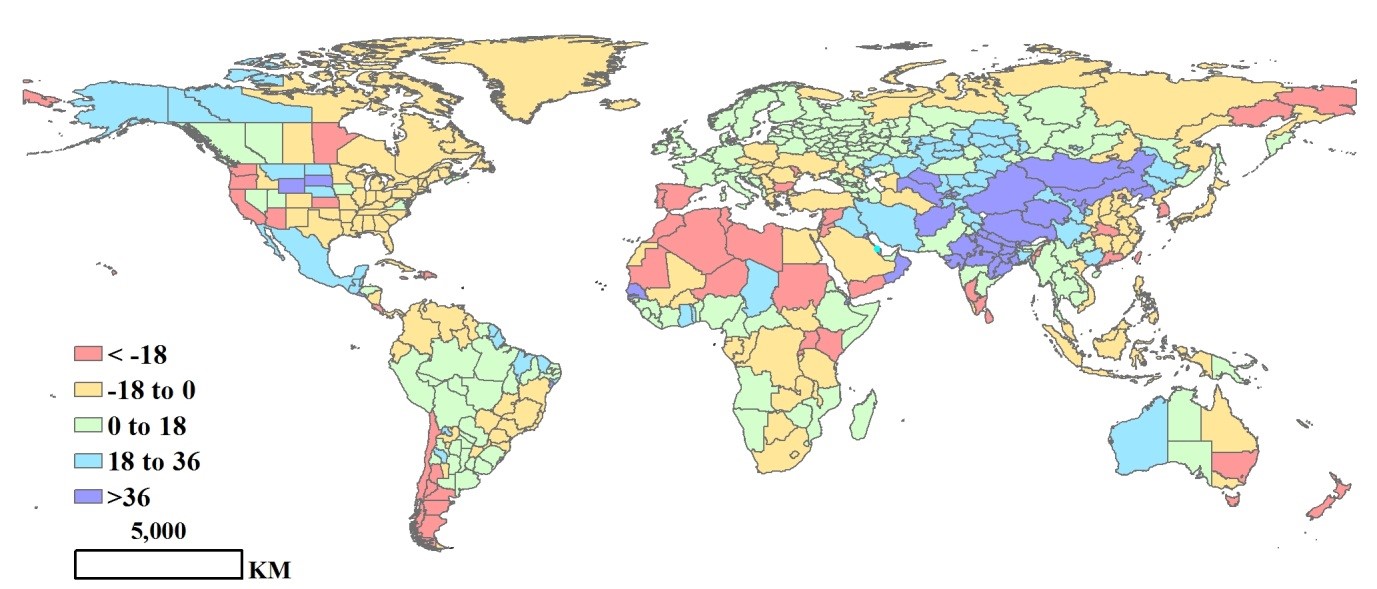
图3.2.全球各国(包括大国的省州级别)最佳植被状况指数
本期CropWatch通报对南半球的阿根廷,巴西和澳大利亚的小麦单产和产量进行估算(表3.2)。三个国家的小麦估算产量都有所增加,阿根廷和澳大利亚的小麦估算产量与2012年产量相比增幅为15%,巴西的小麦估算产量增幅为30%。南非国内资料显示,由于种植小麦获利较低,南非2013年小麦产量下降6%。
表3.2Cropwatch估算的南半球主要国家(澳大利亚,巴西和阿根廷)以及南非小麦产量(2013-2014),单位千吨。
| 小麦 | ||||||
| 单产 | Δ% | 面积 | Δ% | 产量 | Δ% | |
| 澳大利亚 | 2.82 | 8.9 | 3321 | 5.1 | 9356 | 14.1 |
| 巴西 | 1.96 | 11.3 | 13335 | 4.3 | 26181 | 16.5 |
| 阿根廷 | 2.6 | 12.3 | 2212 | 16.9 | 5750 | 31.3 |
| 南非(*) | 1750 | -6 |